УДК
630*453
Д.А. Белов
УРОВЕНЬ
ПОВРЕЖДЕНИЯ И ДОЛЯ ИЗЪЯТИЯ ЛИСТВЫ ЧЛЕНИСТОНОГИМИ ФИЛЛОФАГАМИ В ГОРОДСКИХ
НАСАЖДЕНИЯХ
Значимость комплекса
членистоногих филлофагов как в естественных, так и в
искусственно созданных насаждениях основывается на значимости повреждаемого ими
при питании объекта – листовой пластинки, которая относится к самым
приоритетным функциональным структурам дерева. Площадь поверхности листвы
признается наиболее информативной экологической характеристикой дерева, хотя и
трудоемкой при количественном ее измерении и оценке.
Исследование уровня
потребления первичной зелёной продукции членистоногими филлофагами
является существенным моментом мониторинга лесных и городских фитоценозов,
позволяющим судить об интенсивности происходящих в них процессов. Фоновое
повреждение и фоновое изъятие (освоение) листовой поверхности древесных
растений являются важными характеристиками состояния насаждений и
биоиндикаторами состояния окружающей среды.
При повреждении листвы
видами, присутствующими на своих кормовых породах постоянно и в небольших
количествах, значимых изменений в биоценозе обычно не происходит. Резкое
изменение степени воздействия насекомых на древесное растение характеризует
нарушение баланса взаимоотношений между видами в экосистеме и служит сигналом
происходящих в ней неблагоприятных процессов [14, 15].
Степень влияния филлофагов на древесные растения, в первую очередь, зависит
от размеров изъятия листвы. Большинство авторов в настоящее
время сходится на том, что небольшие потери массы и площади листьев,
соответствующие обычным нормам изъятия (не более 10 – 20 % от общей площади или
массы листьев), не вызывают ощутимых изменений в состоянии растений. Отдельные
исследователи поднимают этот порог до 30 % и даже до 50 % [6, 17].
Установлено, что к концу
вегетационного периода беспозвоночные-филлофаги
обычно повреждают в той или иной степени абсолютное большинство листьев, но
размер изъятия листовой поверхности при этом находится в пределах 30 % [9].
Наносимые членистоногими филлофагами повреждения объединяют в однотипные по своему
характеру и последствиям группы: 1) грубое объедание листвы (хвои), или погрызы, 2) скелетирование
листьев (хвои), 3) сворачивание листьев, 4) минирование листьев (хвои), 5) галлообразование, 6) высасывание соков из тканей листа.
Изучение поврежденности и
динамики изъятия листовой поверхности членистоногими филлофагами
проводилось по известной методике, применяемой рядом исследователей [4, 5, 8,
12, 13, 14 и др.] и модифицированной специалистами кафедры экологии и защиты
леса МГУЛ [3, 7, 11, 16].
В процессе исследований
оценивались два основных показателя для членистоногих филлофагов,
наносящих вышеуказанные типы повреждений: поврежденность листьев (N, %) и
доля изъятия или освоения ими листовой поверхности (D, %), коррелирующая
с массой съеденной листвы. При этом поврежденность являлась количественной
оценкой распространения и встречаемости отдельных видов и групп филлофагов в течение вегетационного периода на изучаемых
видах растений, а доля освоения листовой поверхности – величиной,
характеризующей степень участия видов и комплексов членистоногих филлофагов в потреблении листвы, показателем влияния филлофагов на растения.
Доля освоенной листовой
поверхности (di,
%) определялась для каждого
вида вредителей, для определенного периода наблюдений и вида растения (береза повислая, липа мелколистная, вяз гладкий, рябина
обыкновенная, тополь бальзамический и ива козья). Суммарная доля освоенной
(изъятой) листовой поверхности (D, %) определялась как сумма
долей изъятой листовой поверхности разными видами членистоногих (di) по наносимым ими повреждениям:
D = S di,
где di в свою очередь, определялась как произведение общей
поврежденности листьев (N, %) на
поврежденность листьев данным видом или типом повреждения (ni, %) и на средневзвешенную долю изъятия листовой
поверхности данным видом: (Vi, %):
di = N ´ ni ´ Vi.
Для каждого вида древесных
растений в результате получали общую сумму съеденной или поврежденной массы
листвы по датам учета. По этим данным стало возможным определить долю участия
отдельных видов и комплексов филлофагов в потреблении
листвы и их соотношение, охарактеризовать специфику видового состава филлофагов в различных типах насаждений и его сезонное
изменение, оценить распространение и роль отдельных видов и экологических
комплексов.
Наиболее распространенными
типами повреждений листьев в зелёных насаждениях и городских лесах Москвы
являются грубое объедание, скелетирование,
минирование и галлообразование.
Итоговыми размерами поврежденности
листьев и доли изъятия листовой поверхности были приняты эти показатели в конце
вегетационного сезона.
Данные исследований
поврежденности листьев различных древесных пород филлофагами
(N) за 1992 – 1999 гг. в конце вегетационного сезона, позволяют
утверждать, что у всех шести исследованных пород в конце вегетационного периода
наблюдался высокий уровень поврежденности листвы (от 62 до 100 %). При этом у
березы, вяза, рябины и ивы он определялся поврежденностью листьев в конце
вегетационного периода открыто живущими листогрызущими
насекомыми. У липы в 1992 – 1996 гг. он определялся высокой поврежденностью
листьев листогрызущими насекомыми, а в период 1997 –
1999 гг. – высокой степенью поврежденности листьев минирующими вредителями,
основными из которых были липовая моль-пестрянка (Lithocolletis
issikii Kumata) и в
значительно меньшей степени липовая моль-крошка (Nepticula
tiliae Fr.). У тополя
высокий уровень общей поврежденности листьев определялся, в основном, их
минированием молью-пестрянкой тополевой нижнесторонней
(Lithocolletis populifoliella Tr.) и в меньшей степени
тополевым слоником-блошкой (Rhynchaenus populi F.), а при снижении численности популяций моли-пестрянки
– комплексом скелетирующих листву тополя жуков.
Данные исследований по определению доли изъятия листовой поверхности филлофагами (D) за 1992 – 1999 гг. позволяют утверждать, что этот показатель в конце вегетационного периода для большинства древесных пород не превышал 30 %, и в основном, колебался от 8 – 10 до 15 – 18 %. При этом ведущая роль в изъятии листовой поверхности всегда принадлежала открыто живущим листогрызущим насекомым.
В табл. 1 приведены сводные
данные, полученные на основании материалов о поврежденности и доле изъятия
листовой поверхности указанных ранее шести видов древесных растений, на
основании которых можно сделать вывод о наиболее значимых видах и экологических
комплексах членистоногих, принимавших участие в повреждении этих пород.
В качестве меры их
значимости можно использовать средние и максимальные значения показателя D,
позволяющие сравнить значения этого показателя для разных экологических групп
членистоногих и проследить изменчивость этого показателя по годам.
Таким образом, за
многолетний период наблюдений наибольшие суммарные значения доли изъятия листовой
поверхности (D) наблюдались в 1994 и 1995 гг. за счет высокого
значения этого показателя у минёров и листогрызущих
насекомых. В годы с низкой плотностью популяций минёров относительно высокий
уровень изъятия поддерживался за счет листогрызущих
насекомых и лишь в отдельные годы – за счет галлообразователей
(1992, 1997 гг.).
Приведенные данные позволяют
сделать важные выводы о среднем, минимальном и максимальном фоновых
повреждениях основных лиственных пород членистоногими филлофагами
в условиях городских насаждений и о его флуктуациях. На основании этих материалов возможно выделить виды и комплексы филлофагов, выполняющих наиболее важную роль в урбофитоценозах, о чем можно судить по доле изъятия ими
листвы тех или иных древесных растений.
Полученные в процессе
исследований материалы позволяют также получить для названных выше комплексов и
отдельных видов филлофагов данные об изменчивости уровня
потребления ими листвы за многолетний период, что опосредованно характеризует
динамику их популяций (табл. 2).
Для оценки изменчивости D использовались
его статистические показатели, по которым определялся коэффициент флуктуации –
F(D), характеризующий наблюдаемую кратность увеличения или амплитуду колебания
[18].
Таблица 1
Доля изъятия листовой
поверхности по годам наблюдений (D)
у шести
древесных пород филлофагами за 1992 – 1999 гг., %
|
Годы |
Средний,
минимальный и максимальный уровни изъятия листовой поверхности древесных
растений членистоногими филлофагами (сред. ± m / мин. – макс.) |
|||
|
листогрызу-щими |
минерами |
галлообразователями |
суммарное |
|
|
1992 |
4,27 ± 0,46 1,10 – 8,26 |
4,85 ±3,17 0,01 – 63,02 |
3,89 ± 1,47 0,14 – 21,43 |
12,96 ± 3,09 3,10 – 65,11 |
|
1993 |
5,77 ± 0,83 0,01 – 17,64 |
4,79 ± 3,07 0,01 – 61,19 |
1,84 ± 0,51 0,05 – 8,29 |
12,10 ± 2,80 5,48 – 61,21 |
|
1994 |
5,74 ± 0,76 0,01 – 14,14 |
6,56 ± 4,08 0,03 – 79,04 |
1,78 ± 0,34 0,29 – 6,22 |
14,29 ± 3,69 3,50 – 79,06 |
|
1995 |
8,23 ± 1,25 0,01 – 27,99 |
9,77 ± 5,35 0,01 – 79,72 |
1,34 ± 0,40 0,01 – 3,99 |
19,13 ± 3,65 3,81 – 46,22 |
|
1996 |
6,74 ± 0,84 3,10 – 17,53 |
4,88 ± 2,51 0,01 – 39,92 |
0,87 ± 0,28 0,01 – 10,74 |
12,64 ± 2,61 4,35 – 22,10 |
|
1997 |
6,10 ± 0,70 1,79 – 13,12 |
1,96 ± 0,64 0,01 – 9,98 |
2,39 ± 0,77 0,01 – 4,92 |
11,23 ± 1,16 3,38 – 33,34 |
|
1998 |
5,78 ± 0,83 1,21 – 14,51 |
3,37 ± 1,34 0,03 – 24,86 |
0,75 ± 0,31 0,01 – 1,97 |
11,85 ± 1,38 3,46 – 20,19 |
|
1999 |
7,30 ± 0,90 2,78 – 17,39 |
2,18 ± 0,81 0,15 – 12,86 |
0,45 ± 0,13 0,01 – 1,97 |
10,63 ± 1,01 3,46 – 20,19 |
|
За весь период наблюдений |
6,24 ± 0,42 0,01 – 27,99 |
4,79 ± 0,89 0,01 – 79,92 |
1,66 ± 0,39 0,01 – 21,43 |
13,11 ± 0,95 3,10 – 83,67 |
Он рассчитывался как:
F(D) =
antilog DL,
где DL является логарифмическим
отклонением и определяется следующим образом:
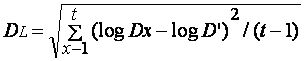 ,
,
где D' – среднегеометрическая
доля изъятия листовой поверхности, вычисляемая как:
![]() ,
,
где D1 ´ D2 ´ ... ´ Dt – измерения
уровня изъятия листовой поверхности в различные моменты времени t.
Значения коэффициента флуктуации
рассчитывались для основных экологических групп филлофагов
в целом и для наиболее часто встречающихся в их составе видов с хорошо
идентифицируемыми видоспецифичными типами повреждений
листьев.
Данные расчета коэффициента
флуктуации F(D) для отдельных комплексов и видов приведены в табл. 2.
Для каждого вида и экологической группы можно указать при этом и размер изъятия ими листвы (D), пропорциональный их уровню численности.
Таблица 2
Статистические значения (D) и его коэффициент флуктуации F(D)
основных экологических групп
и некоторых отдельных видов членистоногих филлофагов
в насаждениях Москвы за восьмилетний период
наблюдений (1992 – 1999 гг.)
|
Комплексы
и виды филлофагов |
Статистистические показатели D |
Коэффициент флуктуации, F |
||
|
средние и ошибка средней, X ± m |
дисперсия, S2 |
показатель изменчивости, V |
||
|
Листогрызущие насекомые |
6,23 ± 0,31 |
14,96 |
62,08 |
1,12 |
|
Скелетирующие листву насекомые |
2,30 ± 0,67 |
25,05 |
217,61 |
28,12 |
|
Минирующие
листву насекомые |
4,79 ± 1,06 |
181,20 |
281,02 |
7,35 |
|
В
том числе: |
||||
|
Моль-пестрянка тополевая нижнесторонняя |
21,41 ± 5,96 |
851,67 |
136,30 |
10,80 |
|
Минирующая тополевая мушка |
0,03 ± 0,025 |
4,49 |
278,81 |
9,60 |
|
Липовая моль-пестрянка |
3,31 ± 0,69 |
23,05 |
145,05 |
2,94 |
|
Липовая моль-крошка |
0,06± 0,05 |
0,016 |
210,82 |
3,70 |
|
Рябиновая извилистая моль-крошка |
0,062 ± 0,033 |
0,009 |
153,01 |
36,43 |
|
Березовая моль-пестрянка |
0,09 ± 0,02 |
0,012 |
121,72 |
4,26 |
|
Березовая чехлоноска |
0,91 ± 0,10 |
0,43 |
72,06 |
3,24 |
|
Ивовая большая моль-пестрянка |
1,27 ± 0,27 |
1,19 |
85,90 |
6,37 |
|
Галлообразователи |
1,66 ± 0,25 |
8,62 |
176,87 |
7,0 |
|
В
том числе: |
||||
|
Грушевый клещик |
1,74 ± 0,73 |
4,24 |
118,34 |
3,03 |
|
Березовый галловый клещик |
0,20 ± 0,10 |
0,076 |
137,84 |
6,19 |
|
Липовый войлочный клещик |
1,65 ± 0,30 |
4,24 |
124,8 |
9,49 |
|
Липовый бородавчатый клещик |
0,40 ± 0,11 |
0,27 |
129,90 |
8,17 |
|
Вязовый мешотчатый клещик |
2,50 ± 0,71 |
12,03 |
138,74 |
7,81 |
|
Ивовый толстостенный пилильщик |
4,37 ± 2,47 |
48,62 |
159,56 |
3,10 |
Значения F(D) указывают на
большое их разнообразие в пределах разных экологических групп филлофагов и у отдельных их видов (от 1,1 до 36,4).
На основании полученных данных о значениях F(D) основные экологические группы филлофагов и отдельных их представителей можно сгруппировать в 5 групп с разным диапазоном колебания этого показателя (табл. 3).
Таблица 3
Сравнение
значений коэффициента флуктуации F(D) с уровнем изъятия
листвы D комплексами и видами членистоногих филлофагов
за
восьмилетний период наблюдений (1992 – 1999 гг.)
|
Значения F(D) |
Комплексы и виды филлофагов |
Значения D, % |
|
1 – 1,5 |
Листогрызущие насекомые |
6,20 |
|
1,5 – 3 |
Липовая
моль-пестрянка |
3,31 |
|
3 – 6 |
Липовая
моль-крошка |
0,06 |
|
Рожковидный
липовый клещик |
0,08 |
|
|
Березовая
моль-пестрянка |
0,09 |
|
|
Березовая
чехлоноска |
0,91 |
|
|
Грушевый клешик |
1,74 |
|
|
Ивовый
толстостенный пилильщик |
4,37 |
|
|
6 – 12 |
>
yle='mso-bidi-font-weight:normal'>Минирующие
листву насекомые |
7,35 |
|
Моль-пестрянка
тополевая |
21,41 |
|
|
Минирующая
тополевая мушка |
0,03 |
|
|
Рябиновая
моль-пестрянка |
0,52 |
|
|
Ивовая
большая моль-пестрянка |
1,27 |
|
|
Галлообразователи |
1,66 |
|
|
Березовый
галловый клещик |
0,20 |
|
|
Липовый
войлочный клещик |
1,65 |
|
|
Вязовый
мешотчатый клещик |
2,50 |
|
|
12 и выше (макс =36,4) |
Скелетирующие листву насекомые |
2,30 |
|
Рябиновая
моль-крошка |
0,07 |
Приведенные в табл. 3 данные позволяют сделать следующие выводы.
Средний уровень потребления
листвы филлофагами (Dсред)
и косвенно связанный с ним средний уровень плотности популяции отдельных
комплексов и видов членистоногих филлофагов в
городских фитоценозах не связан с размахом или амплитудой колебания их
численности, так как значения F(D) практически не зависят от уровня изъятия ими
листвы D.
Так, среди отдельных
комплексов и видов филлофагов с достаточно высокими
значениями коэффициентов флуктуации F(D) – от 6 до 30 и выше, есть
виды и с высокими (тополевая моль-пестрянка), и с очень низкими средними
значениями D (многие галлообразующие клещики и минирующие моли).
Отдельные экологические
группы в целом (листогрызущие, минеры, галлообразователи), за исключением скелетирующих
листву насекомых, в наблюдаемый период времени обладали более стабильным
уровнем потребления листвы, чем отдельные представители этих комплексов.
Принимая во внимание, что уровень потребления листвы членистоногими филлофагами находится в прямой и положительной связи с их уровнем численности, опосредованно характеризуемой показателем F(D), и имея данные о доле изъятия и поврежденности листвы теми или иными видами и комплексами филлофагов и данные об изменчивости этого показателя, можно говорить о характере динамики популяций этих комплексов и видов. При этом следует иметь в виду, что низкий Fсред.(D) = 1,12, вычисленный за 8-летний период наблюдений для группы листогрызущих насекомых в целом свидетельствует лишь о том, что в разнообразных по составу и экологической обстановке насаждениях Москвы фоновое повреждение листвы деревьев листогрызущими насекомыми в наблюдаемый период было более или менее стабильным. Однако очевидно, что колебания уровня плотности отдельных видов этот показатель не отражает.
А.С. Исаев с соавторами (1984),
в соответствии с основными положениями развиваемой ими теории динамики
численности, предлагает разделять насекомых филлофагов
на три группы: эруптивных, продромальных и индифферентных видов.
По отношению к членистоногим
филлофагам в условиях города, Н.К. Белова [1, 2] не
считает возможным выделять среди них типичных эруптивных видов, так как наблюдаемые
в городе вспышки размножения открыто живущих листогрызущих
насекомых протекают иначе, чем в природных условиях. В этом случае предлагается
выделить дополнительную группу членистоногих филлофагов,
занимающих промежуточное положение между продромальными и эруптивными видами.
Для входящих в эту промежуточную группу видов свойственен длительный, почти
перманентно высокий уровень численности и лишь кратковременные периоды ее
понижения, что указывает с одной стороны на благоприятные условия для их
обитания в городе, а с другой – на высокие адаптационные способности к жесткому
антропогенному прессингу. В отличие от видов продромальных, их уровень
численности более высокий, а распространение в насаждениях почти повсеместное. Отличие этих видов от открыто
живущих эруптивных заключается в их скрытом образе жизни (в минах или чехликах)
и нечетко выраженных градационных фазах. В городских насаждениях и
лесопарках Москвы к таким видам Н.К. Белова [1, 2] относит молей-пестрянок –
тополевую нижнестороннюю и липовую, а также лиственичную чехликовую моль.
Анализ
данных, представленных в табл. 2 и 3, позволяет сделать вывод о том, что так
называемые индифферентные виды, средняя многолетняя доля изъятия листвы которыми
невелика (D менее 1 %), способны периодически очень резко увеличивать свою
численность, что определяет для многих из них высокий и очень высокий
показатель F(D), достигающий высоких
значений (более 12). Как правило, такие подъемы численности кратковременны. Однако в
какой-то степени это опровергает утверждение об их стабильной численности. К
индифферентным видам в насаждениях Москвы относятся, за редким исключением,
почти все галлообразователи, многие представители
молей-крошек, жуки семейства трубковертов (Attelabidae),
ряд представителей скелетирующих и грызущих листву
насекомых (например, листовые долгоносики (Phyllobius
sp.), липовый слизистый пилильщик (Caliroa annulipes Kl.), многие виды
листоверток). В комплексе членистоногих филлофагов в
насаждениях Москвы, как и следовало ожидать, по числу видов эта группа
абсолютно преобладает.
К продромальным видам
вредителей в зелёных насаждениях Москвы Н.К. Белова [1, 2] относит березовую чехликовую моль, а также многие виды скелетирующих
и грызущих листву насекомых (многие виды листоедов, листоверток, пилильщиков).
Средний многолетний уровень потребления ими листвы (D) выше, чем у видов первой
группы, но не превышает 4 %, хотя и достигает в отдельные годы значительных
размеров так же, как и значения коэффициента флуктуации. Периоды высокой
численности этих видов кратковременны и колеблются от 1 до 3 лет.
Особо следует рассмотреть
возможность выделения в городских насаждениях так
называемых эруптивных видов. Согласно полученным данным, возможно
отнести ряд видов насекомых в группу эруптивных, которые обладают близкими к
последним особенностями динамики численности и характером очагов массового
размножения. Большая их часть входит в группу листогрызущих
насекомых. Примером таких видов являются непарный шелкопряд и дубовая зеленая
листовертка. Из других групп насекомых можно отнести к ним некоторых минёров –
тополевую моль-пестрянку нижнестороннюю и
лиственничную чехлоноску, которым, несмотря на скрытый образ жизни (у моли) или
полускрытый (у чехлоноски) свойственны выраженные вспышки массового
размножения, напоминающие таковые у эруптивных видов.
Литература
1. Белова Н.К. Массовые виды
филлофагов в зеленых насаждениях г.
Москвы. // Вопросы защиты, охраны леса и озеленения городов. Науч. тр., вып. 224. – М., МЛТИ, 1990. – С. 58 – 64.
2. Белова Н.К. Вредители
городских зеленых насаждений. // Защита растений, № 8. – М. , 1994а. – С. 37 –
38.
3. Белова Н.К., Галасьева Т.В., Куликова Е.Г., Шарапа Т.В. Вредители
растений. // Mетод. указ. –
М., МГУЛ. – 1994б. – 27 с.
4. Богачева И.А. Упрощенный
метод определения доли листовой поверхности, изъятой листогрызущими
насекомыми. // Применение количественных методов в экологии. – Свердловск, УНЦ
АН СССР, 1979. – С. 110 – 116.
5. Богачева И.А.
Абиотические и биоценотические факторы отношений зеленоядных
насекомых с кормовыми растениями в Субарктике. // Ценотические взаимодействия в
тундровых экосистемах. – М., Наука. – 1992. – С. 52 – 59.
6. Богачева И.А. Повреждение
насекомыми листьев березы извилистой в периферической и внутренней частях кроны. // Экология, № 5.– М., 1993.– С. 66 – 71.
7. Галасьева
Т.В. Уровень потребления корма у насекомых-филлофагов
в лиственных насаждениях заповедников. // Растительноядные животные в
биогеоценозах суши. Материалы Всесоюзного совещания. – М., Наука, 1986. – С. 54
– 58.
8. Гуров А.В., Петренко Н.М.
Методы учета трофической активности насекомых-филлофагов
в лесном биогеоценозе. // Лесоведение, № 5. – М., 1988. – С. 15 – 19.
9. Гуров А.В., Петренко
Е.С., Петренко Н.М. Экологическое значение параметров освоения кормовой базы насекомыми-филлофагами. // Лесоведение. 1990. № 6. – С. 49
– 57.
10. Исаев А.С. Хлебопрос Р.Г., Недорезов Л.В., Кондаков Ю.П., Киселев В.В.
Динамика численности лесных насекомых. – Новосибирск, Наука, 1984. – 224 с.
Печатная
версия статьи опубликована:
Экология,
мониторинг и рациональное природопользование // Науч.
тр., вып. 307 (1). – М., МГУЛ, 2001. – С. 50 – 59.