При использовании материалов просьба ссылаться на выходные данные печатного источника или страницу сайта.
Экология заболонников – переносчиков голландской болезни
Е.Г. МОЗОЛЕВСКАЯ, профессор кафедры промышленной экологии и защиты леса МЛТИ, Н.В. КРЫЛОВА, аспирант, Н.К. Белова, старший научный сотрудник, И.Н. ОСИПОВ, студент
Несколько лет назад Московская городская станция защиты зеленых насаждений обнаружила локальные очаги заболонников на ильмовых породах. Появление короедов в городе – сигнал повышенной опасности заселяемым посадкам. Поэтому с 1983 г. наша кафедра по заданию станции приступила к планомерному изучению этих вредителей в городских насаждениях. Установлено, что встречаются преимущественно три вида заболонников – разрушитель (Scolytus scolytus), струйчатый, или вязовый (S. multistriatus), и пигмей (S. pygmaeus). Доля заселенных ими деревьев в локальных очагах на вязовых посадках в среднем равна 6,4 %. Доминирующим видом является струйчатый заболонник, количество заселенных им деревьев (от их общего числа) колеблется в пределах 66 – 95 %, заболонника-разрушителя – 14 – 32 %, пигмея – 5 – 36 %.
Заболонники на ильмовых, известные как активные переносчики голландской болезни, являются объектами всестороннего изучения во многих странах мира. В городах Европы и Северо-Американского континента особенно распространен и вредоносен струйчатый заболонник. У нас заболонники детально исследовались главным образом в лесостепной и степной зонах (Т.М. Гурьянова, 1961; А.Д. Маслов, 1963, 1970, Г.В. Линдеман, 1978). В лесной зоне и в условиях городских посадок их экология в нашей стране практически не изучалась. Заболонники обнаружены преимущественно на деревьях в очагах голландской болезни, реже – на вязах, растущих в неблагоприятных условиях – вдоль трасс с интенсивным движением транспорта, на уплотненных каменистых почвах, в промышленных районах города. Насекомые заселяют ослабленные деревья с изреженной и измельченной или преждевременно желтеющей и опадающей листвой и с различной степенью сухокронности. Они сопутствуют как хронической, так и острой форме проявления голландской болезни, ускоряя темпы усыхания больных деревьев.
Таблица 1
Взаимосвязь размеров жуков-заболонников и ширины их маточных ходов,
толщины луба и диаметра заселяемых деревьев вяза
|
Вид заболонника |
Размер тела жука (мм) |
Ширина маточного хода (мм) |
Толщина луба в верхней границе района (мм) |
Диаметр заселенного ствола, ветви (см) |
|
S. scolytus |
3,5 – 5,5 |
3,5 – 4,5 |
3,5 |
12 – 60 31 |
|
S. multistriatus |
2,0 – 2,5 |
2,0 – 2,3 |
2,0 |
6 – 28 15 |
|
S. pygmaeus |
1,5 – 2,5 |
1,3 – 2,0 |
1,1 |
2 – 12 6 |
Таблица 2
Структура популяции струйчатого заболонника осенью 1986 г. в городских
посадках Москвы
|
Срок массового лета жуков |
Доля особей в популяции (%) |
Соотношение личинок по возрастам, % |
||
|
1 |
2 |
3 |
||
|
Май - июнь |
80,1 |
- |
3,0 |
97,0 |
|
Июль – август |
19,8 |
29,3 |
39,3 |
31,4 |
Жуки втачиваются в толщу коры в местах ее растрескивания или складках, а в зонах тонкой коры – у основания или в развилках ветвей, где она имеет шероховатую структуру. Известно, что толщина коры и луба зависит от диаметра ствола и ветвей вяза. Связь толщины луба (у) и диаметра дерева (х) может быть определена уравнением у = 1,14 + 0,19х. Каждый вид заболонников (в зависимости от величины жуков, глубины и ширины протачиваемых ими ходов) выбирает для поселения участки ствола определенной толщины (табл. 1).
Соотношение видов заболонников в различных посадках определяется условиями, благоприятными для их развития, и прежде всего возрастом и размером деревьев. Так, заболонник-разрушитель доминирует в старых парках, заболонник-пигмей – в молодых посадках, струйчатый – в средневозрастных наиболее распространенных насаждениях вяза. Вертикальное распределение на деревьях зависит от толщины коры: в зонах толстой коры развивается заболонник-разрушитель, средней и переходной – струйчатый, тонкой – пигмей.
Наблюдения показали, что темпы развития этих вредителей не всегда соответствуют темпам усыхания деревьев, они могут или опережать их (при частичном заселении ствола) или отставать от них. На деревьях диаметром свыше 20 см часто наблюдается многолетнее постепенное посекторное заселение ствола заболонниками. В этих случаях на одном и том же дереве могут одновременно развиваться разные стадии или даже разные поколения этих вредителей.
Площадь заселяемой заболонниками боковой поверхности ствола и ветвей более или менее пропорциональна размерам деревьев. Так, при диаметре заселенных струйчатым заболонником стволов от 18 до 25 см площадь зоны поселения изменяется от 1,4 до 7,4 м2. Однако строгой зависимости между диаметром дерева и площадью заселенной боковой поверхности ствола нет, поскольку наблюдается и посекторное частичное заселение предпочитаемых участков стволов медленно усыхающих деревьев. При диаметре дерева на высоте груди 20 – 24 см средняя площадь заселенной струйчатым заболонником боковой поверхности равна 4,3 м2. На ней размещается примерно 1,5 тыс. семей жука, обеспечивающих вылет более 9 тыс. особей молодого поколения.
При планировании и проведении истребительных мероприятий против заболонников необходимо исходить из фенологических особенностей вредителей. В пределах обширного ареала заболонников, совпадающего с ареалом их кормовых пород, в разных природных зонах развивается от одного до четырех поколений.
Продолжительность развития генерации установить трудно из-за растянутого вылета жуков в стациях с различным режимом освещенности деревьев. Так, в условиях Москвы имаго струйчатого заболонника можно встретить с мая по конец вегетационного сезона (рис. 1). Наблюдения за этим вредителем в городских насаждениях, проводившиеся в течение четырех лет, позволили установить преимущественный однолетний цикл его развития с четко выраженным периодом массового лета в мае – июне и зимовкой личинок старшего возраста под корой. Сумма весенних положительных температур до начала массового лета струйчатого заболонника составляла в 1984 г. 476,4°, в 1985 г. – 478° и в 1986 г. – 478,2°. Массовый лет проходил при дневных температурах 18 – 22°. Разница в погодных условиях апреля и мая смещала сроки массового лета в ту или иную сторону на 10 – 15 дней.
К концу июля – началу августе личинки старшего возраста завершают питание и подготавливаются к зимовке, выгрызая колыбельки в толще коры. Весной в этих же колыбельках они окукливаются. В годы с жаркими весенними и летними месяцами часть личинок может окукливаться в первой или второй декаде июля. Тогда вылет молодых жуков, откладка яиц, развитие их и личинок II поколения происходят в конце июля и августе. Это наблюдалось в 1986 г.: примерно пятая часть молодого поколения участвовала во втором лете и развивалась по ускоренному типу. Соотношение двух зимующих поколений и возрастной состав личинок даны в таблице 2.
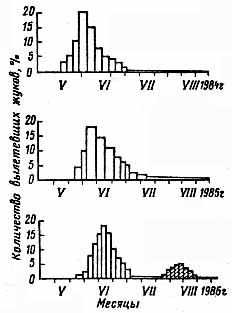
Рис. 1. Динамика вылета струйчатого заболонника в вегетационные сезоны 1984 – 1986 гг.
Таким образом, у струйчатого заболонника в 1986 г. на зимовку ушли личинки старшего возраста 1 поколения и личинки всех трех возрастов II поколения.
Особи разных возрастов существенно различаются не только по размерам, но и по внешнему облику и поведению. Личинки младшего и среднего возраста активно питаются, имеют относительно прозрачные желтовато-розоватые покровы, сквозь которые просвечивает содержимое кишечника; особи старшего возраста после завершения активного питания приобретают молочно-белую окраску и практически неподвижны в своих колыбельках.
Установлено (G. Barson, 1974), что перед зимовкой личинки, завершившие питание, полностью опорожняют пищеварительные органы, теряя в весе почти 1 мг. Это повышает вероятность их выживания. Особи, не закончившие развитие и питание за летний и осенний период и остающиеся зимовать с пищей в кишечнике, погибают.
Интересно отметить, что в Западном полушарии на широте Нью-Йорка, где начало развития струйчатого заболонника совпадает по срокам с началом его развития в Москве, но бывает две генерации в год, наблюдается частичное развитие III поколения, однако его личинки почти полностью погибают во время зимовки (Р.Р. Wallace, R.L. Beard. 1943).
Сроки лета и развития заболонников – разрушителя и пигмея близки к таковым струйчатого заболонника и различаются не более чем на 5 – 7 дней. Весенний вылет жуков и скорость развития молодого поколения у заболонников всех трех видов выше на прогреваемых южных секторах ствола и на хорошо освещенных деревьях.
Таблица 3
Гибель струйчатого заболонника на разных стадиях его развития на дереве
|
Причины смертности |
Количество погибших заболонннков на разных стадиях развития (%) |
|||||
|
яйца |
личинки |
куколки и молодые жуки |
всего за период развития на дереве |
|||
|
до зимовки |
во время зимовки и после нее |
|||||
|
1 возраст |
2 – 3 возраст |
|||||
|
Резистентность дерева |
5,9 |
- |
- |
- |
- |
5,90 |
|
Переувлажнение кормового субстрата |
- |
36,5 |
2,5 |
2,1 |
1,9 |
43,0 |
|
Иссушение кормового субстрата |
- |
- |
- |
0,8 |
0,6 |
1,4 |
|
Паразиты и хищники |
- |
- |
2,2 |
1,2 |
3,2 |
6,6 |
|
Птицы |
- |
- |
1,0 |
1,0 |
- |
2,0 |
|
Болезни |
- |
- |
1,1 |
0,1 |
0,1 |
1,3 |
|
Внутривидовая конкуренция |
- |
- |
0,8 |
- |
- |
0,8 |
|
Неустановленные факторы |
21,3 |
4,3 |
0,8 |
- |
0,6 |
27,0 |
|
Итого |
27,2 |
40,8 |
8,4 |
5,2 |
6,4 |
88,0 |
Таблица 4
Популяционные показатели струйчатого заболонника в зеленых насаждениях города
|
|
Годы развития поколений |
|||
|
1983 – 1984 |
1984 – 1985 |
1985 – 1986 |
1986 |
|
|
Плотность поселения семей (шт. семей/дм2) |
0,3 – 8,5 2,0 |
0,3 – 11,1 3,9 |
0,79 – 6,3 3,0 |
0,9 – 8,8 3,3 |
|
Плотность популяции зимующих личинок (шт./дм2) |
Не учитывали |
0,6 – 122,0 51,2 |
0,4 – 103,4 21,0 |
1,2 – 152,0 51,0 |
|
Плотность популяции жуков молодого поколения (шт./дм2) |
То же |
4,5 – 82,8 44,3 |
8,5 – 73,8 39,8 |
- |
|
Коэффициент размножения |
То же |
9,7 – 14,5 12,1 |
3,1 – 8,9 5,7 |
- |
У имаго после вылета начинается активное дополнительное питание в развилках тонких веточек и в пазухах листовых рубцов. По нашим наблюдениям, на модельных деревьях в среднем на 26,4 % развилок отмечены погрызы жуков, у части развилок от этих мест шли явственные признаки голландской болезни. Дополнительное питание отдельных особей длится 7 – 8 дней, после чего они заселяют деревья, спариваются и откладывают яйца. Заселение деревьев и откладка яиц могут проходить и без дополнительного питания; в наших лабораторных опытах 25 % жуков, не питавшихся; на ветках, заселяли предложенный им кормовой субстрат и давали потомство.
По нашим наблюдениям, самки откладывают яйца в течение 10 – 15 дней, а после завершения яйцекладки большая часть их (90 %) гибнет и остается в ходах.
При заселении повышенно-резистентных деревьев (что обычно наблюдается при высокой численности насекомых) самка заболонников струйчатого и разрушителя, по наблюдениям Г.В. Линдемана (1981), проделывает вместо одного длинного хода два более коротких, обеспечивающих большую вероятность выживания потомства. Заболонник-пигмей изменяет направление и форму начальной части маточного хода с продольной и прямой на крючковатую. Такой ход, перерезающий луб поперек волокон, способствует быстрому подсыханию окружающих тканей и также обеспечивает лучшую выживаемость потомства. При высокой жизнеспособности дерева часть ходов заболонников заливается соком и они совершенно не развиваются; количество таких залитых ходов на отдельных деревьях может быть очень высоким, нами отмечено до 47 %).
Яйца заболонников развиваются около недели. Активное питание и рост личинок продолжаются около месяца. От начала яйцекладки до завершения питания личинок 3-го возраста в естественных условиях проходит около двух месяцев, в лабораторных – 35 – 40 дней. В 1986 г., как уже отмечалось у части молодого поколения заболонников в природных условиях этот срок также сократился до 40 дней.
Мы изучали роль различных причин в смертности струйчатого заболонника в условиях городских посадок. Обобщенные данные для поколения, развивавшегося в 1985 – 1986 гг., приведены в таблице 3.
Как свидетельствуют приведенные в ней данные, резистентность дерева вызывает гибель заболонника лишь на первом этапе его развития (в фазе яйца). Главной причиной смертности вредителя в течение всего периода его жизни на дереве в городских посадках является переувлажнение кормового субстрата. Естественные враги и болезни имеют ограниченное значение. Наибольшая гибель наблюдается, как и следовало ожидать, на ранних этапах развития заболонника, она связана с качеством кормового субстрата.
Некоторые популяционные параметры струйчатого заболонника, позволяющие судить об особенностях его развития в городских посадках, приведены в таблице 4.
Перечисленные показатели характеризуют благоприятные условия развития струйчатого заболонника в зеленых насаждениях города и относительное постоянство условий их обитания и кормообеспеченности. Нам не удалось установить связь темпа размножения (характеризуемого Кр) и плотности поселения семей (р), хотя существует прямая отрицательная зависимость между плотностью популяции и выживаемостью молодого поколения. Абсолютная плотность популяции струйчатого заболонника в его локальных очагах колеблется от нескольких сотен до десятков тысяч жуков, что зависит от количества деревьев на участке и доли ослабленных и больных. При среднем коэффициенте размножения, равном 3,4, во время лета на отдельных участках выходит от 0,6 до 147,2 тыс. молодых жуков.
Коэффициент баланса популяции заболонника на отдельных участках при отсутствии активных мер борьбы равен 1,75, а при интенсивной вырубке заселенных деревьев сокращается до 0,3. Рассчитывается коэффициент баланса популяции как соотношение её абсолютной плотности в текущем и предыдущем годах.
Поскольку почти каждый жук, вылетающий из ствола вяза, пораженного голландской болезнью, является потенциальным переносчиком инфекции, необходима прежде всего своевременная вырубка свежезаселенных деревьев. Основной объем рубок должен выполняться в осенне-зимний и ранневесениий периоды (против зимующих личинок). Необходим строгий квалифицированный надзор за состоянием вязовых посадок и появлением заселенных заболонниками деревьев.
Контроль следует осуществлять на всех участках, где имеются ильмовые посадки. Сроки их осмотра: первая декада мая, первая и вторая декады июня, вторая и третья декады июля, первая декада сентября.
Возможна и химическая защита живых деревьев в локальных очагах заболонников – обработка стволов и толстых ветвей контактными инсектицидами до вылета жуков в середине мая и обработка стволов с зимующими личинками в период их массового окукливания во второй и третьей декадах мая. В городских насаждениях США и ФРГ для уничтожения жуков с высоким эффектом используют феромонные ловушки.
Необходимо тщательно следить за фенологией заболонников и установить окончательно условия, способствующие появлению II генерации этих вредителей. Следует разработать и мероприятия по усилению роли естественных факторов смертности заболонников в их локальных очагах.
Печатная версия опубликована:
Защита растений. – 1987, № 7. – С. 37 – 40.
При использовании материалов просьба ссылаться на выходные данные печатного источника или страницу сайта.