При использовании материалов просьба ссылаться на выходные данные печатного источника или страницу сайта.
УДК 630* 453.78
Н. К.
Белова, канд.
биол. наук, мл.
науч. сотр.
Московский лесотехнический институт
ФАКТОРЫ СМЕРТНОСТИ ТОПОЛЕВОЙ МОЛИ-ПЕСТРЯНКИ
Изменение численности тополевой моли-пестрянки изучали на трех различных участках, выбранных в разных районах Москвы и Подмосковья в течение 1979 – 1981 гг. На каждом из участков были выбраны модельные деревья, которые, по возможности, были типичны для зеленых насаждений участка и удобны для проведения наблюдений. На модельных деревьях с нижних ветвей по окружности кроны брали пробы по 25 – 30 листьев, срывая с каждой ветви по 5 – 7 расположенных рядом листьев.
В статье анализируется влияние различных факторов смертности на развитие тополевой моли и ее выживаемость.
С этой целью проводилось построение таблиц выживания по методике, предложенной Дж. К. Варли с соавторами [2]. Численность популяций тополевой моли в таблицах выживания дается в переводе на 1 лист, то есть по экологической плотности. Основные исследования проводили на фазах яйца, гусеницы и куколки. Попытки учета бабочек на стволах деревьев не дали надежных результатов. Таблицы выживания строились по участкам, а в пределах участков по годам. Обобщенные значения смертности на разных фазах развития по годам и участкам показаны в приводимой ниже таблице.
Из таблицы видно, что значение факторов смертности по годам и участкам обнаруживает значительное сходство. Изменения в количестве яиц, гусениц, куколок и вылетевших бабочек, а также смертность на разных фазах развития в основном были одинаковыми, за редким исключением. Данные таблицы иллюстрирует рис. 1, на котором показано изменение численности тополевой моли по фазам развития на трех участках, выраженное в процентах от начальной численности на один лист, принятое за 100 %.
Значения факторов смертности на разных фазах развития тополевой моли
(по данным таблиц выживания по годам в пределах 3-х участков)
|
Факторы |
Значения факторов смертности, % |
|
|||||||||||
|
смертности |
1979 г. |
1980 г. |
1981 г. |
|
|||||||||
|
|
№ участков |
|
|||||||||||
|
|
1 |
2 |
3 |
Ср. знач. |
1 |
2 |
3 |
Ср. знач. |
1 |
2 |
3 |
Ср. знач. |
|
|
Фаза яйца |
|
||||||||||||
|
Абиотические факторы |
|
|
|
|
14,3 |
15,8 |
17,9 |
16,0 |
|
|
|
|
|
|
Внутривидовая конкуренция |
13,5 |
12,2 |
7,8 |
11,2 |
15,4 |
23,1 |
32,3 |
23,6 |
11,3 |
5,3 |
3,7 |
6,8 |
|
|
Фаза гусеницы |
|||||||||||||
|
Внутривидовая конкуренция |
28,1 |
41,6 |
19,4 |
29,7 |
33,9 |
45,2 |
18,5 |
32,5 |
47,6 |
46.6 |
34,9 |
43,0 |
|
|
Энтомофаги |
14,8 |
15,7 |
13,4 |
14,6 |
13,8 |
18,9 |
8,5 |
13,7 |
15,2 |
9,4 |
8,4 |
11,0 |
|
|
Болезни |
3,2 |
3,7 |
2,5 |
3,1 |
3,4 |
4,9 |
2,8 |
3,7 |
4,8 |
2,8 |
4,2 |
3,9 |
|
|
Неизвестно |
2,8 |
2,4 |
3,2 |
2,8 |
2,6 |
2,2 |
3,8 |
2,9 |
5,6 |
6,8 |
5,4 |
5,9 |
|
|
Фаза куколки |
|||||||||||||
|
Энтомофагн |
6,3 |
5,6 |
6,7 |
6,2 |
11,1 |
12,4 |
11,1 |
11,5 |
4,6 |
8,6 |
14,1 |
9,1 |
|
|
Хищники |
1,2 |
0,9 |
1,0 |
1,0 |
2,6 |
2,8 |
2,7 |
2,7 |
1,9 |
3,5 |
1,7 |
2,4 |
|
|
Болезни |
10,1 |
12,7 |
10,0 |
10,9 |
2,5 |
3,4 |
3,2 |
3,0 |
8,3 |
9,3 |
6,6 |
8,1 |
|
|
Неизвестно |
2,4 |
1,8 |
3,7 |
2.6 |
1,2 |
1,6 |
2,4 |
1,7 |
1,5 |
1,8 |
2,4 |
1,9 |
|
На фазе яйца основным фактором смертности, как и на фазе гусеницы, явилась внутривидовая конкуренция, обусловленная высокой численностью тополевой моли. Смертность при внутривидовой конкуренции зависит от случайного сочетания места и времени кладки яиц. Чем больше случаев неодновременной кладки, тем выше процент смертности. За время исследования на' трех участках в среднем погибло на фазе яйца от внутривидовой конкуренции 13,9 % особей, а на фазе гусеницы 35,1 % особей (минимально 18,9, максимально 47,6 %). Гибель яиц и гусениц происходит в результате неодновременного развития яиц. Запаздывавшие в развитии яйца оказывались на поверхности мин, которые выгрызались ранее вылупившимися гусеницами, и обычно погибали. Даже в тех случаях, когда из таких яиц выводились гусеницы, они, попадая в полость мины более взрослой гусеницы, не могли питаться и погибали.
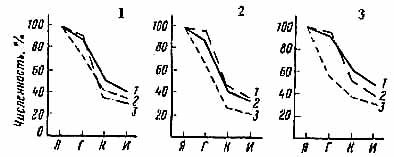
Фазы развития
Рис. 1. Изменение численности тополевой моли пестрянки по фазам и участкам в 1979-1981 гг (Я – яйцо, Г – гусеница, К – куколка, И – имаго, 1 – 1979 г., 2 – 1980 г.; 3 – 1981 г.)
Находящиеся на одном месте гусеницы строго ограничены в пространстве и в пище и не могут выбраться за пределы первоначального заселения. Возникает гибель гусениц, за счет которой остальные получают возможность нормально развиваться. При плотном заселении листа разрастающиеся крупные мины начинают сливаться с мелкими. Маленькие гусеницы, попадая в крупные мины, не находят опоры, пребывают в "подвешенном" состоянии, не могут питаться и погибают.
В 1980 г. наблюдалась значительная смертность от действия абиотических факторов. По трем участкам она составила в среднем 16 %. Весна была холодной, среднемесячная температура составила 8,4° С. Это была самая низкая температура за последние 20 лет. Кладка яиц продолжалась до конца июля, была очень растянута. Часть яиц при этом гибла и разрушалась. Растянутость откладки яиц обусловила неравномерность отрождения гусениц и устройство мин, усилив тем самым смертность от внутривидовой конкуренции.
Смертность от энтомофагов была сравнительно невелика (рис. 2). На фазе гусеницы она составила в среднем 13,1 % (минимум 8,4, максимум 18,9 %), а на фазе куколки в среднем 8,9 % (минимум 4,6, максимум 12,7 %). Видовой состав паразитов был очень небольшим и состоял исключительно из наездников. Всего было обнаружено 7 видов, из которых два определены только до рода: Apanteles bicolor Nees., А. circumseriptus Nees., Sympiesis gordius Walker., S. sericeicornis Nees., Cirrospilus pictus Nees., Chrysocharis sp., Pnigalia sp.
Самым многочисленным был Chrysocharis sp.. Он встречался на всех участках и во все годы и от всех паразитов составлял 39,4 %. Интересно отметить, что В. Полежаев [3] в качестве основного паразита указывает Chrysocharis smirnovi Buk. n. sp., а П.Д. Румянцев [6] – Chrysocharis boops Thorns. Согласно В. Буковскому [1], это один и тот же вид. Вероятно этим же видом является и наш Chrysocharis sp., поскольку он определялся В.А. Тряпицыным, в работах которого С. smirnovi отсутствует. На долю Cirrospilus pictus Nees. и. Pnigalia sp. приходится по 12,7 % от общего числа паразитов. S. gordius распространен повсеместно. От общего количества выведенных паразитов он составлял 15,8 %. Apanteles составили каждый по 3 % от общего числа вылетевших.
Сравнительно небольшую зараженность тополевой моли паразитами наблюдали и другие исследователи. Так, П.Д. Румянцев [6] приводит 5 видов паразитов, из которых до вида определено только два, В. Полежаев [3] – 11, Н.Г. Скопин [7] – более 10 видов наездников Chalcidoidea и Proctotrupoidea, К.Е. Романенко [5] – 4 вида, Фулмек [8] – 32 вида, в том числе 27 видов Chalcididae.
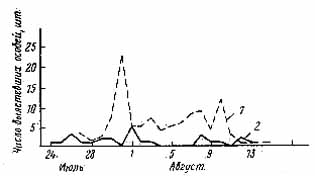
Даты наблюдения
Рис. 2. Динамика выхода имаго и энтомофагов тополевой моли-пестрянки из куколок. 1 – имаго; 2 – энтомофаги
Тахины совершено не были обнаружены в качестве паразитов, что, вероятно, связано с образом жизни гусениц и куколок, защищенных минами.
Хищники и болезни почти отсутствовали и не играли роли как факторы смертности на фазах гусеницы и куколки.
На фазе имаго тополевая моль гибнет при сильных зимних морозах, при весенних заморозках во время лета; во время сильных дождей бабочки смываются со стволов деревьев. Врагами тополевой моли в условиях населенных пунктов являются насекомоядные птицы, уничтожающие бабочек, и паукообразные, охотящиеся за бабочками в трещинах коры деревьев, в дуплах, под отстающей корой.
На всех участках были муравьи Lasius niger L. Они бегали между сидящими на стволах бабочками, но не нападали на них. Зато после дождя муравьи буквально набрасывались на поврежденных и погибших бабочек и большую и #1086;93; часть растаскивали.
По данным таблиц выживания графически выражается только значение показателя К. Он позволяет с большей наглядностью выделить ключевой фактор смертности, имеющий решающее значение в динамике численности изучаемого объекта.
Значение К представляет собой разницу между логарифмами численности популяций предшествующей и последующей стадий развития насекомого.
Так как мы составляли таблицы выживания исходя из предположения, что все виды смертности действовали поочередно, то смертность за поколение можно вычислить как сумму всех смертностей, т. е.:
K = k1 + k2 + k3 + k4 + ... + kn.
На логарифмической шкале изображаются графики всех этих членов, то есть отдельно каждый фактор k и общая смертность К. Каждый из факторов смертности обозначали определенным индексом с коэффициентом k.
Выводы
В итоге можно сказать, что на всех трех участках ключевым фактором, вызывающим изменение численности в популяциях тополевой моли, являлась внутривидовая конкуренция как следствие высокой плотности популяции при ограниченном пространстве и пищевом ресурсе. Однако на участке 1 она выступает как основной фактор на фазе гусеницы, на 2 участке – на фазах яйца и гусеницы и на 3 участке – на фазе яйца, что возможно объясняется расположением участков с юга на север в интервале 25 км.
Это положение хорошо согласуется с работами В. Полежаева [3, 4], который изучил механизмы внутривидовой конкуренции у тополевой моли еще 50 лет назад. К сожалению, эти работы в течение многих лет были преданы забвению.
Литература
1. Буковский В. Новые малоизвестные хальциды (Hymenoptera, Chalcidodea) – Энтомол. обозр, 1938, т 27, № 3 – 4, С. 152 - 171.
2. Варли Дж.К., Градуэлл Дж.Р., Хасселл М.П. Экология популяции насекомых – М.: Колос, 1978, – 222 с.
3. Полежаев В. Борьба за существование у тополевой моли (Lithocolletis populifoliella Тr.). – 3оол. журн., 1934, т. 13, вып. 3. – C. 485 – 505.
4. Полежаев В. Новые данные по экологии тополевой моли (L. populifoliella Тr.). – Вопросы экологии и биоценологии. – Л.: 1939, вып. 4, С. 281 – 289.
5. Романенко К.Е. Нижняя тополевая моль (L. popuhfoliella Tr.) и ее естественные враги в Киргизии //Тр. Киргизской лесн. опытн. станции. Фрунзе, 1959, вып. 2, С. 249 – 257.
6. Румянцев П.Д. Биология тополевой моли в условиях г. Москвы – Зоол. журн., 1934, т. 13, вып. 2, С. 257 – 279.
7. Скопин В.Г. О некоторых пилильщиках и чешуекрылых – первичных вредителях тополей в Юго-Восточном Казахстане. //Уч. записки Казахского университета. – Алма-Ата, 1957, т. 29, С. 103 – 117.
8. Fulmek L. Parasitinsekten der Blattminieren Europas. Den Haag, 1962, 203 s.
Печатная версия опубликована:
Межвузовский сборник научных трудов "Экология и защита леса. Взаимодействие компонентов лесных экосистем" – Л.: ЛТА, 1985. – С. 89 – 93.
При использовании материалов просьба ссылаться на выходные данные печатного источника или страницу сайта.
----------------------------------------------
Свои сообщения оставляйте в Гостевой книге.
-----------------------------------------------
Рубрику ведут к.б.н. Белов Д.А. и к.б.н. Белова Н.К.
“White ant studio” by D.A. Belov and N.K. Belova
A rolling stone gathers no moss. Катящийся камень мхом не обрастет.
_____________________________________________________________
© Белов Д.А., Белова Н.К., 2007 – 2222 г.
Все права на материалы, находящиеся на сайте, охраняются в соответствии с законодательством РФ, в том числе, об авторском праве и смежных правах.
При любом использовании текстовых, аудио-, фото- и видеоматериалов ссылка на сайт обязательна. При полной или частичной перепечатке текстовых материалов в интернете гиперссылка на сайт обязательна.